
![]() Odgovori na
postavljena pitanja - Biologija
Odgovori na
postavljena pitanja - Biologija
411. Molim Vas da napišete nešto o teorijama Hekela, Hadžija i Strinberga o porijeklu višećelijskih organizama. Unaprijed hvala.
U suvremenim teorijama o postanku mnogostaničnih životinja (Metazoa) ističu se tri gledaišta:
J. HADŽI i J. HANSON nosioci su teorije polinuklearnog cilijatnog postanka Metazoa ili polikariontske teorije. Prema njihovoj teoriji prave tkivne mnogostanične životinje (Eumetazoa) nastale su tako što se u njihovih cilijatnih predaka jezgra podijelila više puta, a tijelo je ostalo nepodijeljeno. Oko svake jezgre nalazilo se malo citoplazme te su takve strukture nazvane karionti. Polikariontsko stanje u početku je pokazivalo sincitijalnu građu, a tek kasnije u procesu celularizacije ti su dijelovi odijeljeni staničnom membranom. Time je bio ispunjen uvjet za postanak tipičnog mnogostaničnog oblika. Budući da se u mnogih trepetljikaša može zapaziti tendencija u pravcu razvoja bilateralne simetrije, autori te teorije smatraju da su preci Metazoa bili bilateralno simetrični, a njihov je razvoj tekao preko plošnjaka acelnog (bescrijevnog) tipa, koji sadrži najviše primitivnih oznaka mnogostaničara.
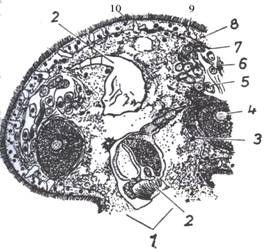 |
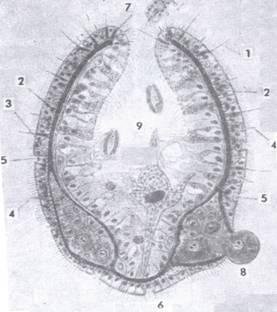
| Slika 1A Dio poprečnog presjeka kroz tijelo bescrijevnog virnjaka (Otocelis sp.) (iz
MATONIČKIN et al.) 1. usta, 2. hrana, 3. jezgra u sincitiju, 4. jaje, 5. spermioblasti, 6. spermiji, 7. bazalna membrana, 8. epiderm, 9. žljezdana stanica, 10. trepetljike |
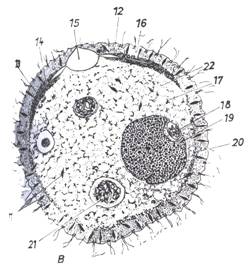 |
| Slika 1B Poprečni presjek kroz tijelo papučice (Paramecium sp.) (iz MATONIČKIN et
al.) 11. trihocisti, 12. pelikula, 13. lipidi, 14. dovodna cijev, 15. stežljivi mjehurići, 16. ektoplazma, 17. mitohondriji, 18. mikronukleus, 19. makronukleus, 20. kapljice masti, 21. hranidbeni mjehurići, 22. entoderm |
Argumenti u prilog ovoj teroriji su sljedeći:
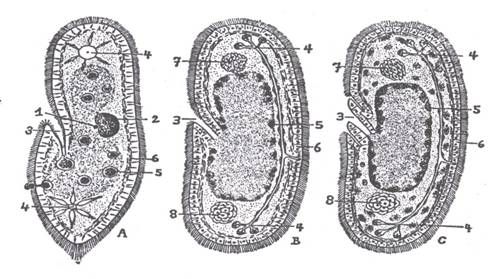 |
| Slika 2 Hadžijeva acelna teorija. Izravno izvođenje bescrijevnog virnjaka (C) iz slobodno
živućeg trepetljikaša (A) preko hipotetskog multikariontskog međuoblika s
početnom celularizacijom (prema Hadžiju) (iz MATONIČKIN et al.) 1. mikronukleus, 2. makronukleus, 3. citofarinks, odnosno usni otvor, 4. stežljivi mjehurić, odnosno protonefridij, 5. hranidbeni mjehurić (entocitijum, odnosno crijevo), 6. pelikula, odnosno epiderm, 7. središnji živčani sustav, 8. položaj gonada |
Polikariontskoj teoriji iznosi se nekoliko prigovora, a to su:
E. HAECKEL postavio je kolonijalnu ili kormijsku teoriju razvoja mnogostaničara. On smatra da su unutar praživotinja neki oblici postali zadružni, oblika šuplje lopte (kao što je danas Volvox). Taj oblik, koji HAECKEL naziva blasteja, bio bi predak svih mnogostaničara (Slika 3). Iz njega se razvio dalji hipotetski oblik jednostavne građe, sastavljen od dva sloja stanica, od kojih je unutrašnji (endoderm) vršio probavnu funkciju. HAECKEL je taj oblik nazvao gastreja (Slika 4). Prema HAECKELU danas živući žarnjaci koji su uglavnom peharasta oblika, a tijelo im je izgrađeno od dva sloja stanica, najsličniji su razvojnom obliku gastreje.
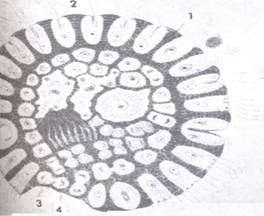 |
| Slika 3 Shematski prikaz hipotetskog filogenetskog stupnja tzv. blasteje (prema Hadžiju)
(iz MATONIČKIN et al.) 1. sluzavi omotač, 2. stanice s bičevima, 3. spolne stanice, 4. oštećeni dio što će ga zacijeliti susjedne stanice iz unutrašnjosti blasteje |
 |
| Slika 4 Shematski prikaz hipotetskog filogenetskog stupnja tzv. gastreje. U donjem
dijelu gastreje između kože i crijeva su gonade (prema Hadžiju) (iz
MATONIČKIN et al.) 1. osjetne stanice s osjetnom dlačicom na slobodnom kraju, 2. indiferentne stanice koje su u bazalnom dijelu epitela, 3. živčane stanice, 4. knidoblasti, 5. ekskrecijske stanice s trepetljikama, 6. aboralni dio tijela, 7. oralni dio tijela, 8. jaje napušta ovarij, 9. crijevna šupljina |
Glavne dokaze za svoju razvojnu teoriju HAECKEL je našao u embrionalnom razvoju mnogostaničnih životinja, jer svaka od njih započinje razvoj od jednostaničnog oplođenog jajeta koje se dijeli u embrionalne stanice. U najjednostavnijim slučajevima zametak prelazi najprije kroz stupanj blastule, zatim gastrule, koje bi trebale odgovarati blasteji, odnosno gastreji. Tu sličnost u razvoju HAECKEL je naglasio u svom poznatom zakonu rekapitulacije filogenije. Organizmi koji stoje blizu hipotetskom obliku blasteje i gastreje postoje i danas među bičašima i žarnjacima. Kod bičaša nalazimo sve prijelaze zadruga s malim brojem stanica, pa do vrsta u kojih je broj stanica vrlo velik. Na primjer, vrsta Gonium sociale ima 4 stanice povezane sluzi, dok Volvox ima 5000 do 20000 stanica.
Osnovni prigovor kormijskoj teoriji je u tome što se u traženju metazojskih predaka ograničila na biljne bičaše kopnenih voda volvokalnog tipa koji imaju celulozne membrane, klorofil i autotrofnu ishranu. Postavljena je i alternativna hipoteza da Metazoa potječu iz neke skupine novonastalih zooflagelata, koji su imali zadružnu organizaciju sličnu volvokalnim biljnim bičašima.
Brazdanje jajeta i tvorba blastule, a zatim gastrule ne moraju biti i najvjerojatnije nisu nikakve rekapitulacije nekadašnjih odraslih stanja. Rano međusobno odjeljivanje blastomera, koje nije opći i obavezni način razvoja, ima svoje fiziološke razloge te je vjerojatno u vezi s potrebom za povećanje površine zbog velike potrebe kisika. Napokon, ni spužve, a još manje knidariji, nisu nikakvi gastreadi, dakle, njihovo tijelo nije dvoslojno. Puštajući askone (najjednostavniji tipovi spužvi) po strani pokazao sam da se knidariji uopće ne smiju uzimati za genetsku vezu između jednostaničara i mnogostaničara. Hidra i drugi hidroidi, tobožnji gastreadi, produkt su natražnog regresivnog razvića. (HADŽI) (Slika 5).
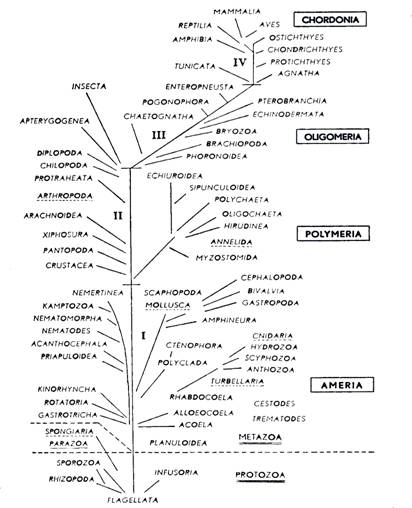 |
| Slika 5. Shema rodoslovnog stabla životinja prema Hadžiju (iz MATONIČKIN et al.) |
Postoje još dvije teorije koje objašnjavaju postanak Metazoa, bliže su Haecklovoj teoriji gastreje, a to su Mečnikova i Jaegerstenova teorija.
MEČNIKOVA teorija zasnovana je na embriološkim podacima, uzimajući u obzir činjenicu da se kod nižih Metazoa gastrulacija odvija na način da stanice vegetativnog pola pojedinačno migriraju u blastocel. Iz tih se stanica razvija endoderm. Te stanice mogu potpuno ispuniti blastocel, pa se u njima naknadno stvori gastrocel. Taj hipotetski oblik MEČNIKOV naziva parenhimula, bio bi sličan planuli - planktonskoj ličinki žarnjaka.
G. JAEGERSTEN (1955) brani Haecklovu teoriju gastreje, ali na drugi način izvodi gastreju iz blasteje. On smatra da se blasteja spustila na dno, gdje je imala bolju mogućnost hranjenja, a zbog kretanja, pri kojem je uvijek jedna strana bila okrenuta prema naprijed i dolje, razvila se u bilateralni oblik, nazvan bilateroblasteja (Slika 6). Zbog fagocitne ishrane na ventralnoj strani životinja se nadvila u luku, te je bilateroblasteja prešla u bilaterogastreju, iz koje su se razvili koralji.
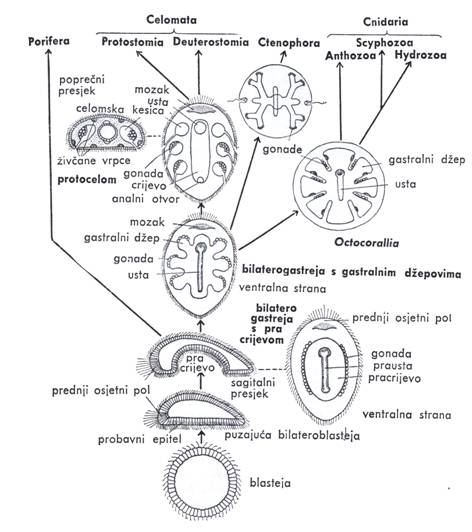 |
| Slika 6. Shema razvoja Metazoa prema Jaegerstenu (iz MATONIČKIN et al.) |
Više autora smatra da Metazoa imaju polifiletski postanak. GREENBERG (1959) sugerira da spužve i žarnjaci, rebraši i plošnjaci imaju nezavisan razvoj od praživotinja. Spužve i žarnjaci razvijali su se iz zadružnih flagelata, a plošnjaci i rebraši od cilijata, a možda i Mesozoa. GREENBERGOVO gledište predstavlja kompromis između kolonijalne i polikariontske teorije, pa je prema tome, ovisno o argumentima koji govore za ili protiv prve dvije teorije.
Literatura:
Dr. sc. Marija Špoljar
viši asistent na Zoologijskom zavodu
Prirodoslovno-matematički fakultet, Zagreb
Povratak na početnu stranicu biologije
© E-škola u suradnji s ![]() om.
om.