
Traheide i traheje su elementi ksilema kroz koje se u višim biljkama na veće udaljenosti prenose voda i u njoj otopljene tvari. Traheje i traheide imaju dimenzije kapilara. To su mrtve, više ili manje produžene stanice s odrvenjelim stijenkama koje ne sadrže citoplazmu što olakšava provođenje vode jer bi citoplazma prijenosu vode pružala veliki otpor. Stijenke elemenata ksilema pojačane su odrvenjelim zadebljanjima koja sprječavaju da se oni stisnu kada u njima pri jakoj transpiraciji vlada podtlak. Ovakva specijalizirana anatomija omogućuje provodnim stanicama ksilema učinkovito provođenje velikih količina vode.
Postoje dva tipa provodnih elemenata u ksilemu: traheide i traheje. Traheide su produžene vretenaste stanice koje su u međusobnom kontaktu preko brojnih pora u bočnim stijenkama. Te su pore mikroskopska područja u kojima nema sekundarne stijenke, a primarna stijenka je tanka i porozna. Pore jedne traheide nalaze se nasuprot pora traheide s kojom graniči. Između takvog para pora nalazi se samo porozni sloj koji se sastoji od dvije primarne stijenke i središnje lamele. Traheje su kraće i šire od traheida i imaju perforirane krajnje stijenke. Te stijenke na svakom kraju stanice čine perforirane ploče. Kao i traheide, i traheje imaju pore na bočnim stijenkama. No, dok su traheide složene u okomite redove koji se preklapaju, traheje se slažu jedna na drugu i čine veću jedinicu koja se naziva cijev. Poprečne stijenke se djelomično ili potpuno otapaju i zato traheje predstavljaju vrlo učinkovit put za vodu. Traheide su prisutne i u kritosjemenjača i u golosjemenjača, dok traheje imaju kritosjemenjače i samo mala skupina golosjemenjača (Gnetales).
Provođenje vode elementima ksilema rezultat je kohezije molekula vode zbog vodikovih veza i adhezije molekula vode na stijenke provodnih elemenata. Rezultat toga je da se stupci vode ne prekidaju čak niti pri jakoj sili sisanja. Hidrostatska napetost unutar ksilemskih elemenata prenosi se na vodu u staničnim stijenkama i na vodu u stabljici. Ako napetost postane prevelika, može doći do stvaranja zračnih mjehurića u provodnim elementima (kavitacija i embolija).
Ukupna površina prereza elemenata za provođenje vode u stabljici biljke po gramu svježe tvari listova koji trebaju biti opskrbljeni vodom ovisi o ekotipu. Biljke suhih staništa imaju više vrijednosti površine prereza nego biljke vlažnih staništa čija je stopa transpiracije niska.
SEKUNDARNE REAKCIJE FOTOSINTEZE
Svi fotosintetski eukarioti od najprimitivnijih alga do najodvedenijih kritosjemenjača reduciraju CO2 do ugljikohidrata istim osnovnim mehanizmom, fotosintetskim C3 redukcijskim ciklusom ugljika koji se naziva se i Calvinovim ciklusom. Ugljik u Calvinov ciklus ulazi u obliku ugljikovog dioksida, a napušta ga u obliku ugljikohidrata. Kao izvor energije koristi se ATP, a kao izvor visokoenergiziranih elektrona potrebnih za redukciju koristi se NADPH. Prvi ugljikohidrat koji nastaje je gliceraldehid-3-fosfat koji sadržava tri C-atoma (zato se biljke koje na taj način vežu ugljikov dioksid naivaju C3 biljkama). Za sintezu jedne molekule tog ugljikohidrata ciklus se mora okrenuti tri puta fiksirajući tri molekule CO2. Pod procesom fiksacije ugljika podrazumijeva se početna ugradnja CO2 u organski spoj.
Ciklus počinje vezanjem molekule ugljikovog dioksida na molekulu ribuloza-1,5- difosfata (RuBP) djelovanjem enzima RuBP-karboksilaze oksigenaze (Rubisco). Nastaje nestabilni spoj sa šest C-atoma koji se odmah razgrađuje u dvije molekule 3-fosfoglicerata koje primaju po jednu fosfatnu skupinu od ATPa i prelaze u 1,3-difosfoglicerat. Visokoenergizirani elektroni iz NADPH reduciraju 1,3-difosfoglicerat u gliceraldehid-3-fosfat koji je ugljikohidrat. Ako je ciklus počeo s tri molekule CO2, dobiva se šest molekula gliceraldehid-3-fosfata. Jedna od njih može napustiti ciklus, a ostalih pet molekula mora se reciklirati kako bi se regenerirale tri molekule RuBP. Za to su potrebne tri molekule ATPa. Za sintezu jedne molekule glukoze potrebne su dvije molekule gliceraldehid-3-fosfata, što znači da je potrebno šest molekula CO2. Da bi se sintetizirala jedna molekula glukoze, u Calvinovom ciklusu se utroši 18 molekula ATP-a i 12 molekula NADPH. ADP i NADP+ koji pri tome nastaju, vraćaju se u svjetlosne reakcije gdje se ponovno fosforiliraju odnosno reduciraju. Korak koji ograničava brzinu Calvinova ciklusa je reakcija karboksilacije RuBP koju katalizira enzim Rubisco. Taj enzim je smješten na površini tilakoidne membrane okrenutoj stromi. Kloroplasti sadrže vrlo mnogo tog enzima, pa on čini više od 16% sveukupnih proteina u kloroplastima i oko 40% ukupnih topljivih proteina u većini listova. U biosferi ga ima više negoli bilo kojeg drugog proteina, a odgovoran je za fiksiranje oko 200 bilijuna tona CO2 godišnje. Godišnje je potrebno oko 107 tona ovog enzima, približno 20 kg po svakom čovjeku.
Zbirna jednadžba sekundarnih reakcija fotosinteze:
6 CO2 +12 H2O +12 NADPH + 18 ATP Ž C6H12O6 + 12 NADP+ + 6 H+ + 18 ADP + 18 PI
Za sintezu jedne molekule glukoze mora se fiksirati šest molekula ugljikovog dioksida te utrošiti 18 molekula ATPa i 12 molekula NADPH. Drugim riječima, po okretaju Calvinovog ciklusa troše se po 2 molekule NADPH i 3 molekule ATPa za svaku fiksiranu molekulu ugljikovog dioksida.
Zbirna jednadžba sekundarnih reakcija fotosinteze jednostavan je sažetak sljedećih reakcija:
(1) 6 ribuloza-1,5-difosfat
+ 6 CO2+ 6 H2O ?Ž 12 (3-fosfoglicerat) + 12 H+
ribuloza-1,5-difosfat
karboksilaza oksigenaza (RUBISCO)
(2) 12 (3-fosfoglicerat)
+ 12 ATP Ž 12 (1,3-difosfoglicerat) + 12 ADP
3-fosfoglicerat kinaza
(3) 12 (1,3-difosfoglicerat)
+ 12 NADPH Ž 12 (gliceraldehid-3-fosfat) + 12 NADP+ +12 Pi
NADP gliceraldehid-3-fosfat
dehidrogenaza
(4)2 (gliceraldehid-3-fosfat)
Ž 2 (dihidroksiaceton-3-fosfat)
trioza-fosfat izomeraza
(5) 2 (gliceraldehid-3-fosfat)
+ 2 (dihidroksiaceton-3-fosfat) Ž 2 (fruktoza-1,6-difosfat)
aldolaza
(6) 2 (fruktoza-1,6-difosfat)
+ 2 H2O Ž 2 (fruktoza-6-fosfat) + 2 Pi
fruktoza-1,6-difosfat
fosfataza
(7) 2 (fruktoza-6-fosfat)
+ 2 (gliceraldehid-3-fosfat) Ž 2 (ksiluloza-5-fosfat) + 2 (eritroza-4-fosfat)
transketolaza
(8) 2 (eritroza-4-fosfat)
+ 2 (dihidroksiaceton-fosfat) Ž 2 (sedoheptuloza-1,7-difosfat)
aldolaza
(9) 2 (sedoheptuloza-1,7-difosfat)
+ 2 H2O Ž 2 (sedoheptuloza-7-fosfat) + 2 Pi
sedoheptuloza-1.7-difosfat
fosfataza
(10) 2 (sedoheptuloza-7-fosfat)
+ 2 (gliceraldehid-3-fosfat) Ž 2 (ksiluloza-5-fosfat) + 2 (riboza-5-fosfat)
transketolaza
(11) 4 (ksiluloza-5-fosfat)
Ž 4 (ribuloza-5-fosfat)
ribuloza-5-fosfat epimeraza
(12) 2 (riboza-5-fosfat)
Ž 2 (ribuloza-5-fosfat)
ribuloza-5-fosfat izomeraza
(13) 6 (ribuloza-5-fosfat)
+ 6 ATP Ž 6 (ribuloza-1,5-difosfat) + 6 ADP + 6 H+
ribuloza-5-fosfat kinaza
3-fosfoglicerat nastaje kao rezultat karboksilacije ribuloza-1,5-difosfata (1) koji se prvo fosforilira do 1,3-difosfoglicerata (2) pomoću ATPa koji potječe iz svjetlosnih reakcija. CO2 se ugrađuje u karboksilnu skupinu "gornje" molekule 3-fosfoglicerata. 1,3-difosfoglicerat se reducira do gliceraldehid-3-fosfata (3) koristeći NADPH iz svjetlosnih reakcija. Ovaj korak katalizira kloroplastni enzim gliceraldehid-3-fosfat dehidrogenaza.
Neprekidna fiksacija CO2 nužno zahtijeva da se akceptor CO2 stalno regenerira. Da bi se spriječilo iscrpljivanje međuspojeva ciklusa, pregrupiranjem ugljika iz 10 molekula trioza-fosfata stvara se šest molekula ribuloza-1,5-difosfata.
Jedna molekula gliceraldehid-3-fosfata pretvara
se u dihidroksiaceton-3-fosfat (4). Tada dolazi
do aldolne kondenzacije dihidroksiaceton-3-fosfata s molekulom gliceraldehid-3-fosfata
(5) i nastaje fruktoza-1,6-difosfat. Taj se
produkt hidrolizira do fruktoza-6-fosfata (6).
2 C-atoma se od fruktoza-6-fosfata prenose preko kofaktora na treću molekulu
gliceraldehid-3-fosfata i nastaje eritroza-4-fosfat i ksiluloza-5-fosfat
(7). Eritroza-4-fosfat se tada kombinira s
molekulom trioza-fosfata (8) i nastaje šećer
sa sedam C-atoma, sedoheptuloza-1,7-difosfat koji se dalje hidrolizira
do sedoheptuloza-7-fosfata (9). On daje jedinicu
od 2 C-atoma petoj molekuli gliceraldehid-3-fosfata i nastaje riboza-5-fosfat
(10). Dvije molekule ksiluloza-5-fosfata (7,
10)
se epimeriziraju i daju ribuloza-5-fosfat (11).
Treća molekula ribuloza-5-fosfata nastaje izomerizacijom riboza-5-fosfata
(12). Konačno se ribuloza-5-fosfat fosforilira
pomoću ATPa i regenerira akceptor CO2, ribuloza-1,5-difosfat
(13).
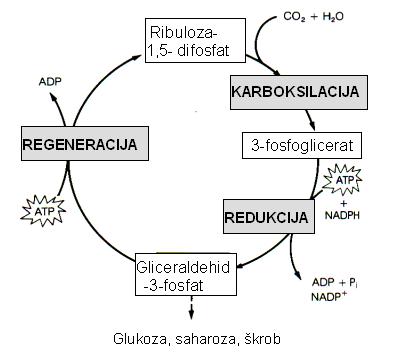
Calvinov ciklus se može podijeliti u tri stadija:
1. karboksilacija akceptora CO2, ribuloza-1,5-difosfata
i stvaranje dvije molekule 3-fosfoglicerata, prvog stabilnog međuspoja
2. redukcija tijekom koje nastaje gliceraldehid-3-fosfat
3. regeneracija akceptora CO2, ribuloza-1,5-difosfata
Prof.dr.sc. Branka Pevalek-Kozlina
Zavod za molekularnu biologiju
Prirodoslovno-matematički fakultet,
Zagreb
bpevalek@public.srce.hr
(C) E-škola u suradnji s ![]() om.
om.